 |
 |
|||
|
||||



| Content of the Workshop (Contribution Dr. Wolter): | ||
| Tuesday, 15.07.2003 | ||
| 11h | "Intact water balance and sustainable management" | |
Wilhelm Ripl and
Klaus-Dieter Wolter
Department of Limnology,
Technische Universität Berlin
Germany
First published as:
Ripl, W. & Wolter, K.-D. (2002): Chapter 11: Ecosystem function and degradation. In: P.J. le B. Williams, D.R. Thomas & C.S. Reynolds (eds.): Phytoplankton productivity. Carbon assimilation in marine and freshwater ecosystems. Blackwell, Oxford: 291-317
for the oral presentation
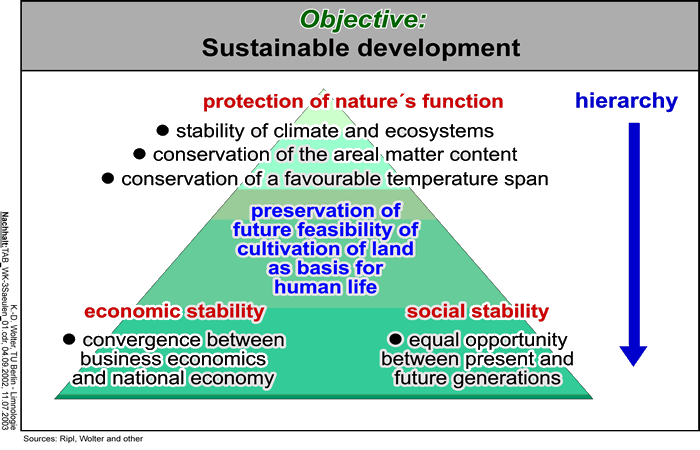
11.1 Introduction and overall aims (Fig. 01)
The sustainability of ecosystem function is now the proper concern of all human society. A stabilised climate, even runoff into surface waters, low material losses, and reliable soil fertility are essential features of an enduring, functional landscape. Through human interference, however, particularly regarding water balance and vegetation, the sustainability of our ecosystems is today very much threatened. We cannot expect to secure improved sustainability without first acquiring a proper understanding of the biospheric functioning and its inherent, self-regenerating processes. What is needed is a coherent and comprehensive recognition of the functions of intact ecosystems, on the one hand, and a knowledge of the responses to anthropogenic impacts that result in their loss of function and degradation, on the other (Fig.2).

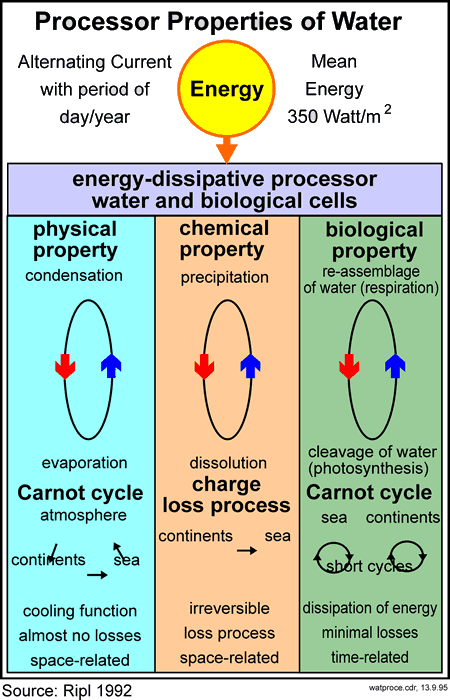
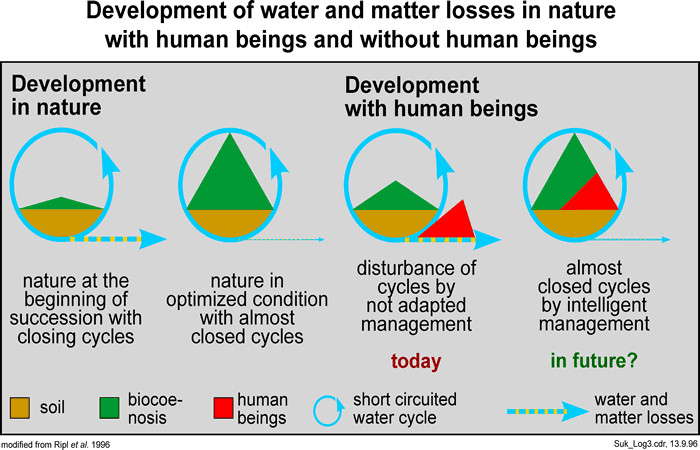
These energy-dissipating processes regulate the ecological dynamics within the earth's biosphere in such a way that the development of natural systems is never allowed to proceed in an undirected or random way. A basic characteristic of unconstrained development in natural systems is the increasing role of cyclical processes while loss processes are correspondingly reduced (cf. Fig. 03, 04, 11). This gives the coincidental increase in the system's sustainability. This might also be viewed as an increase of ecological efficiency (cf. Fig 05, 10), which is applicable at all spatio-temporal levels - from small (microscopic) up to large (whole landscapes). This hierarchy of spatio-temporal levels can be described as a 'fractal' series, the levels being self-similar in their ordering and in their functional tendency towards the proportionate increases of cyclic over loss-making processes.
Energy flow through ecosystems has feedbacks to all spatio-temporal (fractal) levels. The lower levels (smaller in space, localised, shorter in time) always exhibit higher dynamics than upper or higher fractal levels (more space, longer in time). Thus, for example, longer-wave processes, such as forest-generation cycles or the successional development of ecosystems, only appear static from a short-term viewpoint. From a functional viewpoint, however, they should be considered rather as phases, whose harmonic patterns are simply revealed over longer time periods than those of shorter-wave subsystems.

11.2 The efficiency principle
Energy dissipation and efficiency
In this section, we expand the premise that natural systems are energy-dissipative and, thus, self-optimising. As a grounding, the following definitions are fundamental:
- Energy is to the
interaction between two or more objects (e.g. between molecules, or
between the sun and the earth) that results in spatial and/or temporal
change between them (e.g., acceleration or delay in a chemical reaction
or change of temperature).
- Energy dissipation
(the lowering of energy flux) is the process whereby energy gained by
an entity as a pulse (interaction phase and/or from an energy potential)
is released, out of phase, both in time as well as in space. In this
way, the energy pulse is diminished, tending to a dampened or attenuated
value. Thus, the energy potential is reduced, eventually to zero (Ripl
et al., 1996) (Fig
03, 06).
- Water plays a special
significance in global energy dissipation, existing as it does in such
large quantities, in solid, liquid and gaseous form, and showing high
enthalpy leaps between the various phases (see section 11.3) (Fig
03).

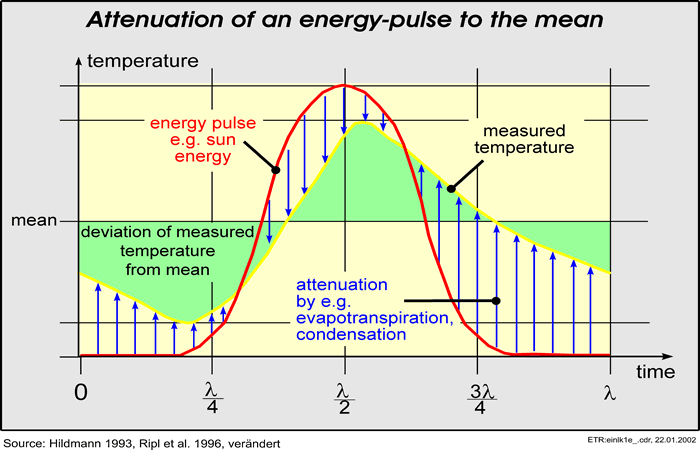
Natural systems that experience unimpaired self-structuring development always approach a condition of lowest energy flux. During this development, energy dissipation is directed increasingly within cyclic processes, so that the proportion of irreversible losses thus being minimised. The system functions more sustainably and its longevity is extended. Given a sufficient period of observation, it has a higher probability of being found than one beset by heavy energy losses (Ripl et al., 1996). In this way, the growth and the attrition of the system become better controlled.
The efficiency of structures may be judged by the extent of their investment in cyclic processes as opposed to loss processes, that is, by their relative closure. It is important, therefore, that delineation of the spatio-temporal units is oriented towards the minimisation of materials transported across their borders. To this end, functional units with minimal openness may be readily recognised at every fractal level:

- atoms with electrons distributed in orbitals about the nucleus;
- molecules with several atomic nuclei and electrons in molecular orbitals;
- molecular aggregates from water (H2O, H+ and OH-), salts and macromolecules, with clusters stabilised by electrical interaction;
- cells with their functional organelles,
- organisms with their different cell aggregates and organs,
- the dissipative ecological unit (DEU; see section 11.3), being the smallest functional unit of a functional ecosystem (after Ripl, 1995a; Ripl et al., 1995),
- ecosystems in aggregate, contributing to global processes.
At maximum stability (permanency), these functional units develop in accord with the efficiency principle. At the levels of atoms, molecules and molecular complexes, they reveal collective pattern - standing waves, oscillations with the same phase, coherency and coupled (cyclic) processes. At the levels of cells, organisms, up to sustainable ecosystems, the functional units are characterised by comparatively stable patterns and extensive cyclic processes, with minimal material leakage to the environment (Fig 11). Thus, in all functional units, independently of their structural level, there is a common pattern. As the proportion of parallel processes (i.e., close, high-feedback production and decomposition) is increased, so the sequential processes at the same level are reduced, so that, overall, energy flux is optimally lowered. Fully functional units dissipate energy almost completely within local cyclic processes. Moreover, irreversibility falls from hierarchically organised levels in a cascade-like fashion.
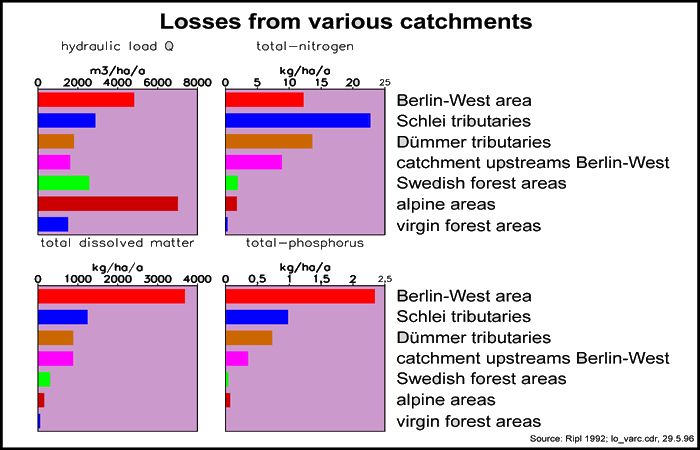
In contrast, degraded functional units are marked by higher randomness in their dynamics, which is accompanied by linear losses (irreversible losses, reduced cyclic processes, more spatio-temporally separated); the longevity of the structure is lowered. Following the second law of thermodynamics (entropy theorem), open systems (i.e. all known systems) are inevitably destined to 'irreversible' breakdown, with unordered movement and structural decay of structure, towards thermodynamic equilibrium. The increase in entropy of a system can be estimated from its irreversible (unusable) energy and/or its material losses (Fig 04). These are also measures of openness; for instance, loss from a hydrological catchment might be quantified in the energy unit, 'proton flow', derived from the total dissolved load of base cations removed in the stream or river, expressed in proton equivalents.
Efficiency of ecosystems
The permanency of natural ecosystems is limited by the available supply of nutrients and minerals in the soil. With a directed water flow, material is transported from the land, into surface waters, and thence to the sea (Fig 04), where the mineral ions thus leached away can only be made available again within geological time periods (i.e. an overall loss process within conceivable time periods). Therefore, only ecosystems that maximise the proportion of matter cycled in relation to that lost irreversibly in the water flow, can be considered sustainable.
In this way, the efficiency of an ecosystem can be measured on the basis of its water and matter balance. Efficiency may be assessed in sub-catchments as well as in entire catchment areas, so long as the minimum relative interchanges with the surroundings are low and the output discharges are known. Two separate measures of efficiency are applicable to ecosystems, - one thermal, the other chemical (Ripl et al., 1995; Hildmann, 1999; Ripl and Hildmann, 2000). Although developed originally for use at the landscape and catchment levels, both coefficients may be applied to other spatio-temporal levels, including those of the organelle, the cell, the organism and the biosphere. Both efficiency coefficients yield results on a scale 0 to 1. Although calculated using different parameters, the two measures of landscape efficiency are directly interlinked - through the deductive reasoning of our concept model - and may therefore be compared directly.

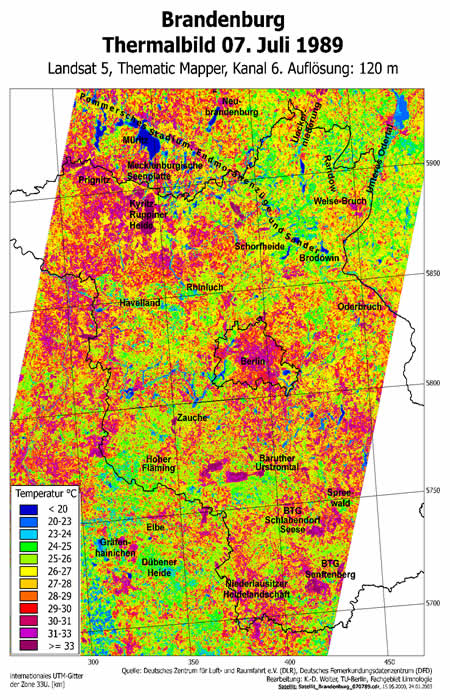
The thermal efficiency (Fig 10) of land systems (the land surface, with its various uses) may be derived from the degree of damping (Fig 06) of the daily, and seasonally-modulated, solar-energy pulse. After a preliminary heuristic phase of investigation, this can be measured from regular satellite observations (using the thermal channel) (Fig 09) on a regional basis.
The chemical efficiency (Fig 09) of an ecosystem may be estimated by the extent of cyclical metabolism that is based on the production of fixed carbon atoms. This chemical efficiency (E) can be calculated as an annual ratio between gross production (P, as a proton yield, 1 mole carbon representing 2 moles H+) and the irreversible loss of cations in the drainage (L, also expressed as 'lost' proton-equivalents as exported base cations). For a given catchment,
Both thermal and chemical measures of landscape efficiency may be used in areas where water is the main medium of the system dynamics; both measures provide valuable inputs to land-use management and planning for sustainability.
Although its fundamental basis is well-known, the efficiency principle has not, to our knowledge, been applied so far to the different fractal levels of various systems. Yet the advantage of this approach lies in its consistent applicability in many other fields of science. By focusing on water and its structuring role as indicators of efficiency, it is possible to deal with the complexity (Fig 03) of entire ecosystems, as well as with the full hierarchy of structural levels. We believe that the method is well-suited to meet the urgent requirement for the determination of manageable development and the management of processes towards sustainability.
For more complete accounts about the functional description of ecosystems and the efficiency principle, as founded on the ETR model, see Ripl (1992, 1995a) and Ripl and Hildmann (2000). The ETR model is based on the dissipative-processor properties of water (Fig 03), the main energy dissipative medium on earth. The model is in accord with the concept of self-organizing dissipative structures (e.g. Prigogine, 1980).
11.3 Ecosystem function at various fractal levels
Dissipative processes at the molecular and pre-cellular level
Liquid water is a fundamental prerequisite for all life processes. In addition, the energy-dissipative reactions in which water is involved, from the interaction between sun and earth down, and its physical and chemical changes of state (e.g. evaporation, dissociation) (Fig 03) make it the most important dynamic medium on the Earth. Together with organic macromolecules, water is the primary constituent of organismic cells (Cope, 1972), where its ionic structure and its mole fractions are predetermined. For instance, the manifold vibrational possibilities of the water molecule and its molecular associations (clusters) give it an unusually high heat capacity. Interactions involving its molecular, atomic and sub-atomic behaviours are subject to unpredictabilities (Heisenberg's uncertainty principle) but the consequences are sufficiently probabilistic for quantitative statements about the functionality of aqueous solutions to be made, even at this level.
Each energy interaction becomes effective and observable in the acceleration or deceleration of matter. By way of electromagnetic radiation and/or mechanical oscillations, relaxing (dissipating) systems interact with absorbing systems, changing their states of motion in the process. In liquids, a higher coherency arises from these localised interactions, harmonic oscillations replacing random movement. Unlike the gas phase, wherein each particle moves in a straight line until it strikes another particle, in liquids, especially in polarized liquids such as water, particles in liquids are substantially coupled. Through electromagnetic interactions, neighbouring particles move together spatially in the same direction, and temporally, with the same frequency. Within such a coherent structure, the probability of a collision with another particle is therefore much reduced. Only within the proximity of phase boundaries, or other structuring of system material, does some modulation of the oscillation process occur. Here, transitions between the coherent vibrations of clusters and the vibrations of ions and macromolecules lead to a local increase in the likely reactions. As a function of amplitude of the oscillations, there results an intensified dissociation of the water. This applies equally to living cells: the same efficiency principle, demonstrable at the molecular level, is repeated and amplified at successive fractal levels.
In aqueous solutions, oscillation patterns are modified substantially by the presence of ions and macromolecules. In the close vicinity of a positively- or negatively-charged ion, the polarised structure (dipoles) of the water molecules causes their relatively rigid alignment. Through changes in bond lengths and angles, local increases in molecular densities also occur in the vicinity of hydrophilic ions or molecules (Gutmann, 1978). In the immediate environment of ions, therefore, the aqueous solution becomes strongly structured. The electrical field associated with individual water molecules is substituted by mutual hydrogen bonding and developing hydrate coverings and clusters. At the interface between these organised and coherently oscillating clusters, there is a zone of 'distortion' within which fewer hydrogen bonds arise, through the disturbance of orientation by the electrical field. Because of the lowered, 'looser' structuring, this zone is particularly fluid. Within a salt solution, discretely distributed differentiated areas arise from the structured changes in the distribution pattern of charge and in the vibration pattern. Thus, due to the oscillation of water molecules, charge density distributions within the salt solution are altered. These then have a sorting effect on other molecules, including macromolecules. Such sorting of polymerisation products would have been critical in the evolution of life: the present-day function of macromolecules in living cells must have evolved by a similar mechanism.
The reactivity of water encompasses acid-base reactions (H+ and OH-), and thus the dissolution and precipitation of salts, as well as the redox reactions (reduction and oxidation); alternately splitting and joining of water (in, respectively, photosynthesis and respiration) (Fig 03). Reactivity is particularly high at phase boundary surfaces (liquids/solids, gas/liquids, etc.). Owing to differences in the natural frequencies of the materials interfaced at these state-change boundaries, individual water molecules experience a form of mechanical stress, which leads to a higher dissociation rate. Accordingly, the electrical energetics (electron distribution in different chemical bonds) and the mechanical energetics (vibrations of atomic nuclei, atoms and molecules) become separated. In this way, the point of reduction of a proton by an electron is reached. By chemical convention, this point is assigned, under normal conditions, the pH value of zero. Thus, the probability of finding mono-atomic hydrogen (i.e., pH=0) reaches 1. In correspondingly small, delimited areas, the pH value is precisely steered in space and time, by way of this modulated oscillation of water molecules. The reactivity of the water and/or radicals, based as it is on the mobility of electrons and protons, subject to the abundances of carbon dioxide, organic molecules (e.g. enzymes) or metals (Cunningham et al., 1987), determines its chemical and biochemical reactions. Far from being an inert solvent, water is an essential component in the structure, transport and reactivity of life processes. Phase boundaries represent the preferential reaction sites for all biochemical life processes.
On the same conservative principle, energy received by a system (e.g. a light energy pulse - a photon) must be dissipated. As a consequence of its function, the system partitions the pulse among chemical energy and heat.
Conceptual models of elementary, energy-dissipative life processes
Because of its rather unique characteristics, water probably also played an important role during the emergence of life on earth. Energetically, the processes described above, at the molecular level of water, explain satisfactorily the emergence of photosynthesis. Conceptually, life processes owe their origins from the structure of water, as outlined below.
Light - the electromagnetic interaction between the earth and the sun - leads to an excitation of terrestrial electrons. Modulation of coherent cluster oscillations is achieved by transformation of absorbed radiation into mechanical movement of the absorbing molecules. The quantity of such molecules is regulated by equilibrium between the rates of formation and decay and, therefore, is controlled by the coherency of molecule movement. Bicarbonate ions or carbon dioxide are integrated into the water structures described above, according to their mole fractions. Reduction to a carbohydrate radical (- CH2O) will take place at reaction centres activated through resonance by mechanical energy, when pH=0 (formation of a hydrogen radical, as above) is reached. Such spatially minute and temporally short-lived reaction centres, it may be assumed, are not in areas of 'free' water, but rather in the proximity of phase boundaries (e.g. sub-microscopic fissures in rocks, or light absorbing, macromolecular modulating oscillations). The selection of the best suited macromolecules is controlled by evolution according to the efficiency principle.This explanation of the origin of photosynthesis would also account for the fact that the oxygen so liberated originates from the water and not from the carbon dioxide.
According to the efficiency principle, the carbohydrate radicals formed should be transported directly to sites with low energy state, where their far-from-thermodynamic-equilibrium state would be most stable (i.e. where coherent oscillations of molecules and ions is lowest). When meeting with other radicals, carbohydrates (e.g. glucose) are formed and further polymerisation leads to the formation of, for example, starch and cellulose, further lowering their kinetic energy. Since the vibrations of macromolecules are of longer wavelengths (because of their higher molecular weight) than are those of water molecules, a discretely distributed, non-Gaussian spatial structure develops at the level of molecular associations. Likewise, their dynamics are not normally distributed, resulting in the sorting and selection of the structures and processes involved (i.e. the probability of reaction) through resonance coupling. This process is sustained until the rates of assembly and disassembly of these substances achieve a dynamic equilibrium. Thus, a higher overall efficiency for the structure is obtained when the openness of the subsystem is lowered and its material losses are minimised.
A further possibility for increasing the efficiency of the overall structure would be the establishment of better matter cycles at the next higher fractal level (in the present case, in the cell association). This 'jump' is the mainspring for self-organisation, bifurcation and for the development towards gradually more complex and larger-scale structures represented by the evolution of organisms and the development of ecosystems on earth. It must be assumed that the necessary reactions could not have arisen simultaneously, neither were they normal in distribution but, equally, they must have appeared very early in the development of living systems. This means, in turn, that a thermodynamically-based selection procedure arose in the lowest levels of molecular self-organisation. The system had turned from that of 'Boltzmann space' (random, chance distributions of processes) to that of 'Hilbert space', in which chance events were eliminated by process coupling - and therefore a substantially higher degree of coherency and patterns would have arisen.
Today, it is generally accepted that self-structuring and self-organising dissipative structures arose early in geological time and that, eventually, these led to the emergence of ever more complex structures and life itself (e.g. Lehninger, 1978; Ebeling and Feistel, 1982). Teleological and comprehensive explanations for the self-organising phenomena of nature (e.g. Resch and Gutmann, 1987) have so far been classified as philosophical and of having no scientific basis. On the other hand, the stabilisation of simple biochemical cycles (e.g. hypercycles after Eigen and Schuster 1979) and smaller biochemically-active particles (e.g. 'coaservates' and 'micro-spheres') could hardly be explained with the usual selection criteria that had been considered up to that time (Ebeling and Feistel, 1982). The suggestion would therefore seem to be to demand that the more traditional explanation for development - by way of random selection and trial and error - should give way to the outcome of the efficiency principle operating at a series of small to large scales. Thus, chemical and biological evolution would be governed by the same functional principle. Structures that are optimised in standing-wave patterns, or spatio-temporally closed cyclic processes, survive for longer than structures subjected to heavy losses. The latter are structures exhibiting a more random dissipation of energy with a higher proportion of irreversible processes. Accordingly, structures with a higher efficiency and higher stability with regard to these cyclic processes (efficiency optimisation), will almost certainly be selected for over longer periods of time (see also Eigen and Winkler, 1993). In this way, biochemical evolution and the development of ecosystems can be just as readily explained - as a thermodynamic necessity (Ripl and Hildmann, 1997).
Use of energetic patterns by cells and organisms
At the level of cells and organisms, the efficiency principle operates in the organisation of material flows through low-loss, coupled, cyclic structures. The cell components (structured water, organic and inorganic substances contained in water, as well as the organelles, membranes and DNA which they constitute) are arranged in, to a large extent, stationary positions. Nevertheless, they are closely involved in cyclic operational sequences. The cell membrane governs the exchange of material with the outside environment and in so doing minimises it. The materials needed in cell metabolism (water, ions, low-molecular organic substances and gases) are transported selectively at rates regulated by the cell membrane. Since all biochemical reactions and transported materials are carried in an aqueous medium enriched with organic and inorganic components, organisms could be just as well regarded as 'structure-optimised water' (Ling, 1984; sea also Cope, 1972). Controlled energy dissipation keeps the processes far from chemical equilibrium, while their efficiency is optimised harmonically with regard to the dynamic surroundings. Coherent, resonant, vibration patterns and short-circuited material cycles are generated.
For single-celled organisms, the efficiency principle already applies: the more effectively the cell can turn over the material running through structured cyclic processes and the fewer losses it makes, then the more stable and thus more survivable it becomes. Thus, the intake of food needed for operating processes is minimised. In multicelled organisms, this efficiency criterion is supplemented by the synergetic division of labour between cells and cell tissue - and by the organism's internal transport systems (e.g. phloem and xylem in plants, blood and lymph circulatory systems in animals). In addition, multicellular organisms minimise a part of their irreversible losses by cycling material internally.
Once again, use of the energy-dissipative efficiency concept for understanding processes at the level of organisms proves helpful. In the distribution of an energy potential, organisms always possess several degrees of freedom for their use of this energy (i.e. to partition it into chemical reactions or allow a rise in temperature). Examples of energy use by organisms include: absorbed electromagnetic radiation (light) and the mechanical transport of reactants. Some organisms use the energetics of their own environment for themselves - and thus operate vital functions with smaller amounts of internally-produced energy (i.e. from NADH + H+ , NADPH + H+ or ATP), rather than by the exclusive use of any one of these energy forms.
Some examples of these functions, hitherto almost ignored, are noted. They include strategic mechanisms to overcome supply deficits, such as CO2 deficiency or a lack of light. They also include feedback processes for rectifying an unwanted surplus, such as of oxygen evolved in excess under bright light. In this manner, the possibilities for organisms to attenuate an existing energy pulse to a mean value are greatly extended; matter cycles are more readily short-circuited and closed.
The Dissipative Ecological Unit (DEU) - the interdependence of organisms
At the next higher fractal level - between organisms - close coupling between two organisms can become more effective, as is seen, for example, in symbiotic nitrogen fixation or in the use of algal exudates by bacteria. However, the more familiar interactions among organisms cover such familiar topics as competition, predation and coexistence. We wish to progress our consideration of the fractal hierarchy by following the same principles and adopting the Dissipative Ecological Unit (DEU) as the next fractal level of organismic interaction.
The idea of a minimum unit ecosystem was developed during the "Stör" river project, carried out in northern Germany, and conceived the context of the ETR model (Ripl, 1992; Ripl, 1995a; Ripl et al., 1995; Ripl and Hildmann, 2000). Here again, efficiency may be understood as the criterion for selection - favouring closure by cyclic processes, minimisation of those that are irreversible and the greatest retention of resources. Higher efficiency in a compound dissipative structure means that it can spread itself more rapidly than an adjacent structure with lower efficiency. It is more stable and it is more sustainable.
At the level of ecosystems, the DEU represents the smallest unit in which evolution and succession takes place and for which a thermodynamic efficiency can be determined. The structure comprises five functional components, capable of mutual coupling in such a way that water and material cycles can be internalised and, to a large extent, closed. The five components are as follows:
1. Primary Producers, having the dual function of manufacturing material and providing the energy needed for all heterotrophic structures, and of pumping the second functional component - water - as coolant, transport and reaction medium, through the process of evapotranspiration.
2. Water - through evapotranspiration - provides a feedback control of production.
3. The detritus buffer - the store of organic matter, the fine capillary structure of which controls the entrance of air (including oxygen) for the fourth component.
4. The decomposers (consisting of bacteria and fungi). Nutrients and minerals are recovered from the detrital store of material and energy when water-saturation is sufficiently alleviated for the microbial mineralisation to proceed. Thus, the store can be used efficiently with a low level of losses. The first four components are subject to a fifth, processing component, regulating energy dissipation through localised structures incurring minimised losses.
5. This fith processing component is the food chain, the network of consumers consisting of lower and higher fauna, creating space by the management of primary producers and decomposers and, thus, also maintaining the important process of reproduction and increasing overall dissipative efficiency.
Within the DEU, there is regulation through a potentially perfect feedback, wherein the 'bottom-up' and 'top-down' controls collapse into one another, in favour of a basic 'control loop'. Its stability and durability may be assessed from the temporal efficiency of local resource use. This control loop implies that there is a management span, in which management by the food chain is regulating energy efficiency in such a way that the daily energy pulses, for example, are smoothed to give an increased efficiency of energy use. With resource cycling at a maximum, residual energy potentials can be better deployed. Expressed in ecological terms, relating to the processes of production and respiration, this means the maximisation of gross productivity but with a reduction in net productivity towards zero. Energy pulses are more completely dissipated and energy flux is maximally attenuated. In a typical DEU, attenuation of the energy flux proceeds by way of: (1) evaporation of water; (2) development of biomass; (3) development of short (and thus more closed) evaporation-condensation cycles, (4) with internal matter cycles; and (5) by the warming of the soil.
As an example, we can consider the spatio-temporal components of processes comprising a functional DEU. We may imagine an experiment with two treatments, both including water and identical quantities of algal and bacterial cells. In the first treatment, algae and bacteria are randomly distributed throughout the water. Switching a light source on and off results in fluctuations in, for example, the levels of oxygen, dissolved phosphate, pH, etc. The exchange of materials will always be made by the relatively long distance of open water separating algae and bacteria with fluctuations of the above determinands arising as a function of the light/dark phases.
In the second treatment, the same quantity of algae and bacteria are closely juxtapositioned, interconnected and interdependent as they might be in periphyton. With the lighting switched on, the oxygen produced by the algae can be respired immediately by the bacteria in a short, localised cycle. Moreover, nutrients and carbon dioxide set free by the bacteria may be taken up substantially more rapidly by the algae. For the same given conversion of material, the second treatment experiences distinctly smaller fluctuations in the determinands than would be measureable in the first treatment. For the same energy flux, the more interconnected community in the second experimental treatment has operated more efficiently and has minimised its material losses.
From this simple reconstruction of a DEU, it may be hypothesised that, through recursive feedbacks among all components over time, harmonic coherent patterns can develop, involving appropriate, mid-range abundances of functionally necessary organisms (primary producers, decomposers and consumers). Moreover, a well-structured DEU should be capable of surviving a pulse of external, disruptive forcing. Provided their frequency is low, such forcing events may be absorbed (the system is resilient). However, disruptive forcing at high frequencies or intensity may exceed the resilience of the DEU, leading directly to its degradation.
The efficiency of a DEU can be determined from the rate that matter is lost relative to the incoming energy flux. Rising matter losses are indicative of poorer attenuation of the energy flowing through the DEU. Conversely, DEUs having high efficiency are longer-lasting than DEUs of a lower efficiency: available minerals (base cations) tend to be retained more effectively than they are in lower-efficiency structures. The most effective DEUs will have all five components well-represented and functioning well.
An indication of the comparative efficiencies of various habitats (DEUs) can be shown by continuous, automatic temperature records, lower-efficiency units showing wider temperature fluctuations than those in units with higher efficiencies (Ripl et al., 1995).
Ecosystem efficiency of landscapes
How completely a landscape dissipates its daily input of solar energy is a measure of the efficiency of the entirety of its ecosystem(s). The extent to which the energy income to a delimited land surface (e.g. a catchment, subcatchment, or the domain of aggregated DEUs) and over a given time period (e.g. a year, or the complete lifecycle of longer-lasting primary producers such as forest trees) is dissipated is a measure of the efficiency of the whole ecosystem. However, ecosystem efficiency, too, has inherent limits. If the energy pulse of a certain frequency were fully damped within the given boundaries of a system (suppressing all spatio-temporal variability in energy flow and material supply, or matter budget, across the given surface), then matter-cycling would be perfect and loss-free, and the efficiency of the system would equal unity. The second law of thermodynamics, however, precludes this limit ever being reached (perpetual motion machines do not exist) and a certain degree of sub-system openness persists.
The notion of measurable ecosystem efficiency can be applied only to defined partially closed spatio-temporal systems, like a DEU or larger unit, such as a sub-catchment. Where the limits cannot be so defined, the functional interrelationships (such as matter flow) will be arbitrarily separable and the system's efficiency will be unquantifiable. The larger the unit considered, the easier the system delineation becomes. Estimates of matter losses - from a single plant, from a forest stand, from a subcatchment or a larger river basin - are self-evidently scale-dependent. According to this logic, the time-scale for assessment needs to be set appropriately to accommodate the generation times of its characteristic components (that of a tree in some instances) when interpreting ecosystem processes.
11.4 Development of ecosystems - self-organisation and degradation
In this section, we seek to demonstrate, by reference to particular types of ecosystem, how the efficiency that is transmitted through the fractal sequence of biological function may be assessed. For a series of functional biospheric compartments, we contrast two extremes of organisation:
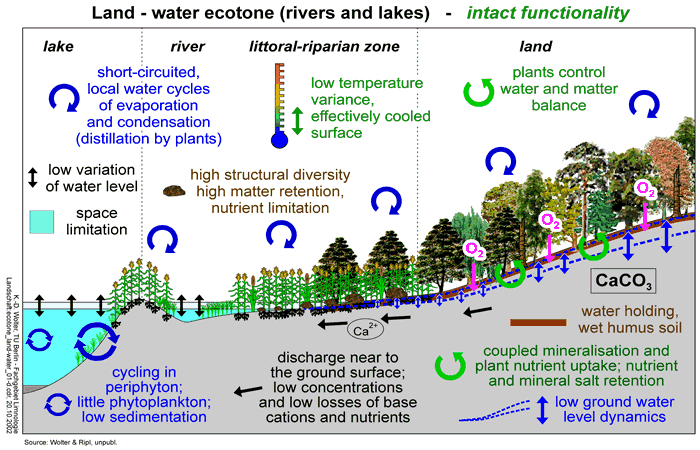
- intact, enduring systems that are self-structured and self-optimised (Fig 11, 13),
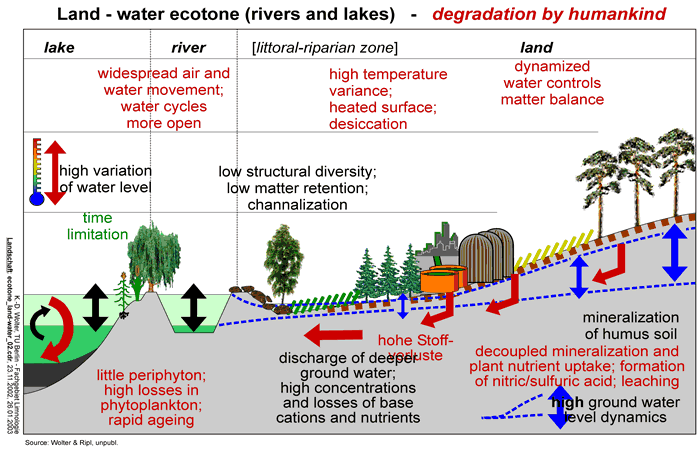
- transient, degraded systems that are losing structure (Fig 12).
There are, in reality, many intermediate conditions but the 'intact' and 'transient' categories are adequate to delimit the range of ecosystem function. They also make it possible to see generalised targets for the reorganisation and restoration of ecosystems and the achievement of sustainability.
As discussed in the preceding sections, all ecosystems intercept energy that is essentially pulsed and material resources (water, resources) that are essentially also in flux. Intact ecosystems optimise the dissipation of the energy and minimise material losses. Transient or degraded systems fail to retain energy or material resources and are ultimately unsustainable. Self-organisation works towards making open systems more intact, by damping fluctuations in energy income, minimising temperature fluctuations and water loss and by increasing the cycling of resources.
Under each of the following section headings, we contrast the functions of intact and transient compartments of the biosphere by reference to the ways in which water transformations are regulated. Thus, optimising functions characterise the self-organisation of intact systems; degrading functions distinguish transient systems. Differences are emphasised in the boxed tabulations. The text provides some quantitative amplification where appropriate.
Atmosphere
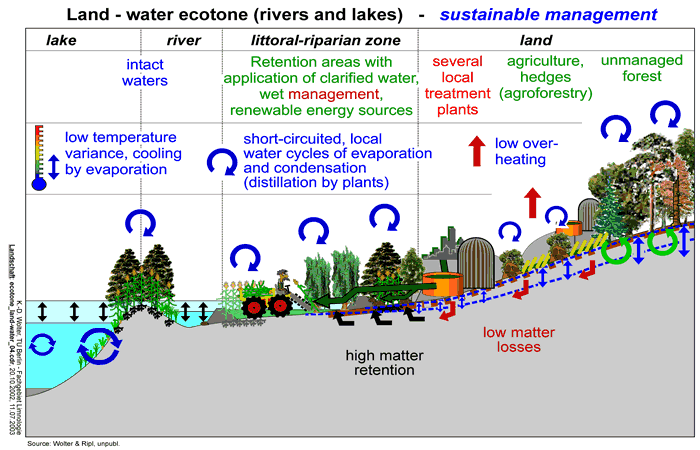
The atmosphere is the capacious medium for the long-range cycling of water between the oceanic global material sink and the continents. The hydraulic discharge from the land masses averages ~ 250 mm a-1 in total, against a mean precipitation over the continents of ~ 670 mm a-1 (Zachmann, 1976). The difference (420 mm a-1) corresponds to the conventional estimate of the mean evaporation from the land to the air. However, this computation obscures the exchanges due to high-frequency, local-scale cycling by evaporation and condensation (e.g. dew and hoar-frost formation on cooled surfaces) which make up a substantial part of the water cycle. Exchanges of water exceeding the estimated precipitation but lost invisibly in a larger evaporation are indicative of a short-circuited water circulation in an intact atmosphere.
The consequences for optimisation of this compartment at the continental scale are noted in Box 11.1. The near-ground cooling by evaporation enhances the presence of vapour near the ground surface where it provides a 'thermal-protection filter' against accelerated solar heating.
Atmospheric degradation
may result from destruction of the permanent vegetation covering and impairment
of the water relations. Reduction in evaporation over large areas allows
wider diel and seasonal variations in air temperatures and larger differences
between the drier, over-heated surfaces on the one hand and, on the other
hand the better-cooled, damper areas (high spatial temperature gradients)
(Fig 07,
09). Near-ground thermostatic buffering is impaired by the loss
of wet surfaces, while intensified periods of dryness lead to apparent
increases in warming over large areas. Dusts, possibly toxin-laden, carbon
dioxide, nitrogen oxides, methane, ozone formed near the surface and chlorofluorocarbons
(CFCs) are more readily transported into the upper atmosphere. Their interference
with global heat exchanges is already well-known. Meanwhile, the reduction
in evaporative cooling and the absorption of heat further degrades the
atmosphere. The increasing reduction of the surface area that retains
its dissipative evaporation-condensation cycles seems to be a contributory
factor in modern climate change.
Box 11.1 Self-organisation and degradation in the atmospheric water compartmentOptimising functions
· Frequency of evaporation-condensation in small-scale cycles is increased.
· Capacity of local diel exchanges is raised.
· Large-scale, non-local water cycling is reduced.
· Dust minimised.Degrading functions
· Temperature damping weakened.
· Thermostatic buffering of near-ground water vapour is lost.
· Heating, drying, air movements, dust generation in large-scale cycles are intensified.
Vegetation
A well-developed, tree-dominated vegetation intercepts a large proportion of the incoming solar energy which is invested in the further production of biomass. An intact vegetation is in close feedback with the local water regime of its location (Box 11.2). During the course of succession, the vegetation develops a humus cover that improves water retention and, through evapotranspiration, further damps the daily energy pulse. The cooler surface has a feedback in favouring more condensation of water. Localised cycling turns the well-vegetated landscape into a dynamic water reservoir. Added to storage capacity of the soil's humus layer, runoff to rivers is progressively moderated. Water retention also contributes to lower redox levels in the litter layer, whence spontaneous mineralisation losses are minimised. The slow release of ions is more effectively cycled back to the rooting zone. In this way, decomposition and production become closely coupled (in parallel), benefiting the retention of water, matter and limiting nutrients. (Fig 11).
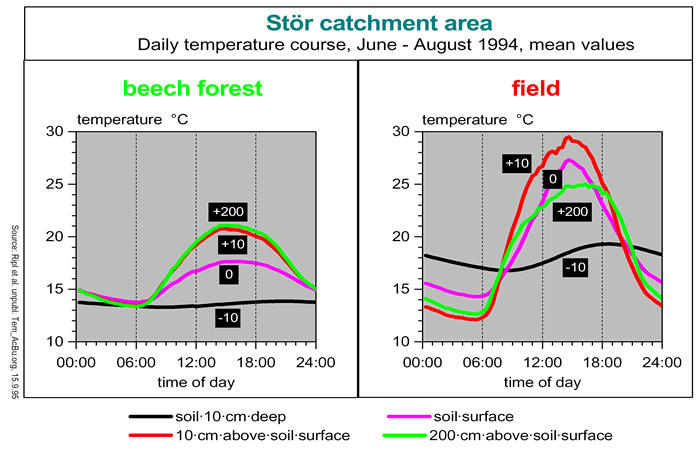
Vegetation is degraded principally through reduction or removal of natural, diverse woodland cover, its replacement with agriculture or commercial monocultures and through accelerated drainage of wetlands. The risk of desiccation is increased, followed by humus oxidation and the loss of temperature damping. On sunny days, unregulated heating may raise the temperatures of vegetation-free surfaces (arable fields, city streets) by more than 10°C above the comparable forest (Fig 07, 09) (Ripl et al., 1995; Hildmann, 1999; Ripl & Hildmann, 2000). The control of short-circuited circulation exerted by vegetation is lost (Fig 11).
Box 11.2 Self-organisation and degradation in the vegetation water compartment
Optimising functions
· Plant succession maximises solar energy harvest.
· Stores of water and humus increased.
· Temperature fluctuations are minimised.
· Dissolution and irreversible losses of materials are minimised.
· Lowered redox levels resists local loss of minerals.
· Retention and internal cycling of resources are increased.
Degrading functions
· Water exchanges are less controlled: inundation and desiccation are more common.
· Humus is increasingly oxidised.
· Mineral salts and nutrients leaked to drainage or removed irreversibly in agricultural crop.
Soil
Unhindered development of vegetation is accompanied by the accumulation of a humus layer in the upper horizon of the soil. Its presence cuts evaporative water losses from below, so helping to maintain the water-retentive capacity and moderate soil temperature (Box 11.3). In heavily saturated soils, the nitrates and sulphates formed in soil water near the surface are rapidly diminished by denitrification and desulphurisation. Liberated hydroxyl ions precipitate base cations that are retained as hydroxides. The resistance to rainwater that humus layers provide encourages lateral water movement (runoff), also minimising the leaching of nutrients beyond the root zone. With the arrangement of the processes of matter production and decomposition in parallel, long-range translocation and misalignment of material with the water flow are avoided (Fig 11).
Box 11.3 Self-organisation and degradation in the soil-water compartment
Optimising functions
· Moisture content and water content are moderated by accumulating humus.
· Mineralisation, especially denitrification and desulphurisation, is retarded.
· Root-zone translocation and resource misalignment is resisted.
· Runoff discharges are moderated.
· Ecotone gradients (with atmospere, vegetation and groundwater) are flatter, more interconnectedness of organisms is facilitated.Degrading functions
· Drainage of agricultural, forested and wetland areas diminishes temperature damping.
· Increase in unsaturated zone and mineralisation with associated acid formation impairs feedback inhibiting material flow.
· Groundwater abstraction increases desiccation and permeability of upper soils.
Ascribing such importance to soil water level, we prefer a more robust distinction than is usually made by hydrologists, between the dynamic, near-surface soil water and the deeper store below the water table and where dynamic exchanges are generally weak. The true groundwater of intact soils supports a small, relatively even discharge with a low mineral and nutrient content.
At the soil surface, with optimal cooling of the surface, the gradients of important parameters (e.g. moisture content, temperature, substrate) are reduced at ecotone boundaries. At major habitat boundaries with sharp gradients in material turnover (e.g. the land-water ecotone), an optimisation and stronger interconnectedness of organisms develops through the process of succession and leads to a highly diverse and efficient community.
With the intensification and industrialisation of agriculture, many soils have suffered degradation. Drainage and continuous cultivation usually result in the increasing mineralisation of soil humus and in the loss of water-retention capacity. The dynamics of near-surface water are accelerated, while deeper groundwater is left unprotected. Gradients are sharpened and some microhabitats are diminished. On the broader scale, water relations are greatly modified (Fig 12).
The symptoms, summarised in Box 11.3, are those of much accelerated water exchanges. Episodes of drying and water logging of the humus-depleted soil also accelerate material losses, respectively through mineralisation and leaching. Structure and, eventually, fertility is lost.
Before the industrial era, agricultural production was largely confined to drained, nutrient-rich, organic soils (e.g. of calcareous fens). Lowering of the level of water saturation would lead to aeration and oxidation of organic matter and to wider varition in soil moisture and temperature. With improved drainage, the formerly closed water and material cycles would be opened. With the oxidation of reduced nitrogen and sulphur compounds, strong acids (nitric and sulphuric acid) would be more prevalent. In an analogous manner, the higher partial pressure of CO2, resulting from the decomposition of organic substances would contribute to acidification with the formation of carbonic acid.
Since the industrial era, these leaching processes have accelerated with the raising of net productivity, itself made possible by the increased use of additional (mostly fossil and nuclear) energy. Today, the main cause of degradation is, ironically, is 'land improvement' for agriculture and forestry (soil drainage, deep ploughing), multiplied by expansion in land area affected. Soils cease to act an intact interface between bedrock and vegetation. Rather, they are reduced to an industrial substratum for the maximisation of agricultural production. The plants (in particular, trees used in commercial forestry) have had to adjust to the increased losses of base cations (Ca, Mg, K) and phosphate by producing acids in their root zones, which, in turn, leads to greater acidification of drainage water. Apart from the incidence of highly acidic atmospheric depositions, this process has provided a significant contribution to the acidification of soils and water (Fig 12).
Material losses (Fig 04) have greatly accelerated as a consequence of this industrialisation of the landscape. Dissolved loads to river drainage have increased by factors of between 5 and 100 (Ripl, 1995a). In terms of calcium, magnesium and potassium alone, German streams transport cations from the land at rates of between 185 and 367 kg ha-1a-1 (Ripl et al., 1996; Hildmann, 1999).
Running waters
An intact catchment area mediates time-equalised output discharges, in which nutrient and mineral ions are simultaneously dilute. With unidirectional drainage, material losses tend to be greater from upper catchments than from the lowlands: the naturally structured catchment imposes an increasing downstream trend to capture and retain matter and minerals transported in surface or soil-water flows.
Through their structure (morphology, colonisation by organisms) and discharges, rivers mirror the water and matter balances of their catchment areas. From an intact catchment, generating an optimally equalised discharge throughout the year (low amplitude between high and low water) and shedding only minor leachate loads, the structure of the drainage network should also be expected to be as stable, diverse and self-organising as possible. In fact, natural river flows conspicuously fashion their channels and adjacent riparian features which, together with the persistent sorting of bedload materials throughout the stream, contribute to a a maximisation of flow-resistance and hydraulic retention. Along these self-modified channels, flowing water is subjected to numerous small-scale accelerations and delays. Examples are provided by the gradients of velocity over single pebbles through to the acceleration and delay zones associated with meandering. The transport capacity of flowing water decreases in favour of small-scale flow resistance. The small-scale flow velocity gradients favour not only a high variety in sorted material, and thus of ecological niches, but at the same time the development of a diversified community. A highly-structured and mainly stable running water is the result - i.e. a long-lasting functioning stream or river (Fig 11).
Box 11.4 Self-organisation and degradation in running waters
Optimising functions
· Intact catchments damp fluctuations in water flow.
· Catchment structuring captures and retains matter against unidirectional drainage.
· Channel formation and riparian development resist flow and favour retention of water and materials.
· Bankside ecotones are flattened, habitat breadth is increased.Degrading functions
· Piecemeal catchment management increases amplitude of flow, increased soil erosion and loss of fertility.
· Canalisation and rivetting to "improve" drainage impoverishes fluvial habitats and diminishes riparian function.
· Severity of flood and drought incidents become more severe.
Sustainable rivers carry only small amounts of entrained material. Depending on their position within the catchment area and their geomorphology, stream banks tend to be rather gently sloped, so that small increases in discharge are sufficient to inundate the riparian zone. In flat and broad streams, where water is disrupted in areas of high flow resistance (riffles), there is enhanced gas exchange with the atmosphere, bringing stabiliisation of O2 and CO2 levels.
We deduce that rivers also go through conspicuous self-optimisation process, favouring the establishment of stable, part-closed biological systems, tending to retain materials rather than void them in the flow (cf. Box 11.4 above).
Degradation of flowing waters features opposite trends. The present-day, near arbitrary arrangement of land usage, coupled with the intensive interference to the natural passage of water (drainage, abstraction, irrigation, centralised sewage treatment and waste-water disposal, channel canalisation) have mostly amplified fluctuations in runoff and accelerated the leaching of matter from the land. Moreover, agricultural tillage is often carried out to the stream edge, encouraging erosional soil loss, which is then countered by revetting or other reinforcement. River beds are narrowed and deepened. Natural riparian storages are removed. It is of little wonder that floods are more common or more severe, that plant life and fluvial microhabitats have been lost and that rivers carry more particulate matter, more leachate and more nutrients than heretofore (Fig 12).
Standing Waters
Lakes and other impoundments, particularly standing waters with shorter residence times, are subordinate to the intactness and degradation of the water and matter balance within their catchments. During development of the landscape (such as in the post-glacial period) terrestrial matter losses are reduced and the trophic state of the receiving lakes decreases (Digerfeldt, 1972). Overloading the metabolic capacity of the lake with the losses of matter from the catchment will lead to degradation to a lower functional efficiency. By contrast, intact lakes have the metabolic capacity to be able to process their material and nutrient loads in an efficient manner. There are intergrades between the intact and the transient lake type: those whose condition would be described, on the conventional trophic scale, as mesotrophic, approximate to the point of transition between the two types, although any perception of metabolic functionality based only on nutrients and production hardly takes full account of the internal dynamics of the lake system (Fig 11).
The concept of intactness and transience adopted here applies to all kinds of lakes (from arctic to tropical, high to low altitude, very small to very large). Behavioural differences are often greatest between deep and shallow lakes. (Limnologically-speaking, shallow lakes are defined as having a maximum depth which is less than 4 % of the diameter of a circle equal in area to that of the lake (Wetzel, 1983). Almost all lakes are plainly open systems - outflowing volumes are balanced against inflowing hydraulic discharge but some or all of the particulate income is retained through sedimentation. They can develop a long-lasting functionality if the catchment loads are relatively modest and if permanent losses of organic matter to outflow and to the sediments are minimised.
In intact lakes, the catchment load is collected and retained largely in the transition zone between land and water, within the subdivision of the system (mesohabitat, or biocoenosis) contributed by littoral vegetation with a well-established network of highly-interconnected organisms, including the periphyton (biofilm) and a well-developed, near self-sufficient food web. Subject to physical constraints, the continuous supply of colonisable material facilitates the expansion of the littoral vegetation, with a positive feedback in terms of matter retention (Björk, 1988; Brinck et al., 1988,). A well-developed littoral structure also buffers waves, so that dead and decomposing material accumulates until it is successively amenable to invasion by swamp and marsh plants and, eventually, by terrestrial vegetation. In this case, the lake becomes overgrown starting from the banks, pointing to a littoral type of sedimentation (Lundqvist, 1927).
The open-water pelagic zone of the intact lake functions on very dilute nutrient base, in part because much of the small phosphorus load is rendered unavailable by a stoichiometric excess of P-precipitating materials (including of iron, aluminium and calcium). The impoverished nutrient capacity can support only a very small producer biomass (phytoplankton) but this nevertheless sequesters a significant fraction of the energy flux through the rapid microbial cycling of the carbon fixed. The production to respiration (P/R) quotient in the intact pelagial is close to 1. The sedimentary output from the pelagic comprises little more than refractory organic substances and biominerals (silica, carbonate), accumulating at less than 1 mm a-1. Conversely, the poverty of the organic carbon supply offers a limited energy source and restricts the habitat suitability of the deep benthos to consumers of detritus and microbial decomposers. Redox levels remain positive. Ageing and silting-up of the lake are slow and longevity (sustainability) is maximised.
Development of an intact lake is constrained by the seasonal pattern of hydraulic and nutrient loading. The theoretical retention time of a lake has a fundamental influence on its overall functioning. In a lake with minimal external loading, closely-connected coupling among the organisms are strongly favoured and within which organisms are increasingly able to regulate their own surroundings (nutrient content, organic substances, final metabolites, light availability, etc. Thus, intact lakes supplied by intact catchments are durable, sustainable structures and they are reckoned to have high value for species diversity (Ripl, 1995b).
The symptoms of transience in degraded standing waters (summarised in Box 11.5) provide stark contrasts. Degradation may have several drivers but the major generic cause is catchment degradation. Following the deforestation that proceeded from neolithic times (approximately 5000 BP) and, especially, since the start of industrialisation of agriculture the exploitation of non-renewable energy sources (in just the last 150 years), material cycles in the catchment areas of lakes have been increasingly opened. As the first interface is at the surface, the reactivity of the lake is much influenced by the ratio of catchment area to lake surface (Ohle, 1971). Some early responses may be restricted to certain parts of the lake - and be exploited by newly-introduced plants and animals (e.g. Canadian waterweed, Elodea canadensis; grass carp, Ctenopharyngodon idella) - but symptoms soon proliferate through the communities of the lake. However, it is not the organisms that are necessarily problematic so much as the modified environment provided by enhanced deliveries of nutrients, organic substances and metabolites. Mostly, the problems arise from elevated metabolic capacity and biological overproduction in the water, spilling over in increased sedimentation and ageing. This process is known as eutrophication.
Box 11.5 Self-organisation and degradation in standing waters
Optimising functions
· Although subordinate to their catchment areas, lakes act as capacitators of hydrologic and material inflows, damping reactivity to fluctuating inputs.
· Mesohabitats and biocoenoses are stabilised with internal recycling of resources.
· Succession in littoral increases material retention and flattens ecotones between water and land surfaces.
· In open (pelagic) waters, sparse resources become segregated and stored in the sediments; phosphorus is precipitated with iron or calcium present in stoichiometric excess.
· Modest organic recruitment to sediments offer limited microbial energy sources.Degrading functions
· Accelerated catchment losses stress the capacity of internal lacustrine processes.
· Material overloads shortcut internal recycles, deregulating pelagic production
and increasing the incidence of phytoplankton blooms.
· Increased turbidity diminishes submerged littoral growth.
· Phosphorus-binding capacity is stoichiometrically exceeded, providing a positive productive feedback.
· Sedimentary flux of organic material is increased; mineralisation depletes oxidative capacity and low redox of sediments and deep waters constrain habitat availability and function.
Excess nutrient availability obviates the close, interconnected resource regulation and releases the environmental constraints on fast-growing, opportunist organisms (r-strategists). For instance, if larger biomasses of phytoplankton and periphyton can be supported, then the light and, in turn, the space available to macrophytic vegetation is compressed and the diversity of the littoral components is diminished and their function is impaired. The stoichiometric capacity of P-binding minerals is saturated and phosphorus ceases to provide a key limitation on phytoplankton production.
While degradation in shallow lakes (i.e., dominated by littoral) is presented first as a 'bottom-up' effect of enhanced nutrient loading, it is also possible to produce analogous effects from 'top-down' adjustments to the multiple feedback provided by fish-stock levels. Increased stocking of white fish (carp, roach, etc.) or the increased harvesting of predatory fish may alter the structure of the food web, reducing the zooplankton and increasing the phytoplankton (Hrbácek, 1994).
Meanwhile, production (P) in the pelagic zone, increases relative to respiration (R) and sedimentary output of organic carbon (P/R >1) is enhanced. The oxidative capacity of the deep-water and the sediments is increasingly challenged, and organic sediments accumulate more rapidly than 1 mm a-1. However, it remains difficult for benthic consumers of detritus and microbial decomposers to exploit the carbon richness because of a lack of oxidant. Specialist anaerobes, such as sulphate-reducing bacteria and methanogens contribute to the degradation of more easily degradable organic substances. The hydrogen sulphide thus formed leads to 'rapid lake-ageing' (Ohle, 1953, 1954): trivalent iron (Fe3+) is reduced (to Fe2+), liberating small amounts of phosphorus bound to the iron into the water by the mechanism described by Mortimer (1941/1942). However, for the release of large quantities of phosphate from the sediment, divalent iron must be scavenged by free sulphide and precipitated as FeS. Thus, P-binding capacity is effectively lost (Ripl, 1978; Ripl & Lindmark, 1979). In this way, the internal metabolism of lakes is matched to processes at the sediment-water boundary layer: degradation of sediment metabolism and the subsequent release of phosphate exerts a positive feedback on production in the pelagic. With the sedimentation of excessive quantities of organic substances, the automatic control loop is opened and substituted by a transient metabolism (Fig 12).
The process of eutrophication is explained conventionally in terms of the increased inputs of phosphorus into surface waters. The basis of this explanatory model was developed by Vollenweider (e.g. 1971), from the statistical analysis of data from a series of large lakes. Originally, most of the additional nutrient was supposed to emanate from point sources, in particular, from sewage purification plants. As a consequence, many sewage purification plants in industrialised countries have been equipped with phosphorus-removal plants, which lowered the phosphorus content in waste-water discharges to far below 100 µg P dm-3. However, only in few cases has a return to oligotrophic conditions been achieved. Diffuse, non-point sources and the release of phosphorus from lake sediments seemingly continue to supply sufficient phosphorus to maintain the prevailing eutrophic conditions. The role of diffuse sources of P from inappropriate, maladjusted land management has increased and has compensated, to a large degree, for the measures undertaken by water management to control P.
Whether the nutrients originate from diffuse or point sources is of secondary importance to the view that, in the end, both are symptomatic of landscape degradation consequential upon several interrelated pressures - groundwater abstraction, food production and waste-water collection. 'Point sources' are merely the foci of an altogether degraded, inefficient water and material balance (see Ripl, 1992).
Nevertheless, eutrophy represents only a temporary phase in the development of a lake. A similar condition would have been encountered during the brief period following the last glaciation when a poorly-vegetated landscape suffered with high matter losses. Eutrophic lakes persist only until either the landscape is fully leached or the lake becomes finally terrestrialised. In contrast to a stable system, degraded lakes, together with their catchments, are characterised by a tendency towards accelerated change. This can lead not only to their rapidly silting-up, but also to their acidification, if base cations are also depleted from their catchments.
Nutrients and minerals at the surface of a catchment represent limited resources. If they are leached because of human interference, then both the vegetation and associated water circulation degrades. The processes of temperature-equalisation from functioning evaporation and condensation cycles, a substantial mechanism for normal temperature distribution on earth, is thereby lost. As a function of the continentality of climate, the continued existence of lakes as surface waters is then put into doubt. Under these aspects, furthermore, the eutrophication of lakes can be brought into connection with the desertification process (collapse of the water and matter balance within a landscape), through which, also, many larger lakes in continental areas are already subject to regression (e.g. Aral Sea).
11.5 Summary and conclusions (Fig 15)
An integrative understanding of the coherency in the process we call 'nature' is indispensable for sustainable management. In the present text, we have tried to construct, on the basis of the dissipative water cycle, an integral deductive model ('Leitbild') for ecosystems. An integrative approach - the Energy-Transport-Reaction (ETR) model - is used to describe the essential processes and functions in ecosystems. Therein, the daily pulse of solar energy is dampened through the dissipative medium of earth - water interaction. In constrained systems, unhindered in their development, external loss processes (away from system borders) are replaced by internal cycles. Losses from systems may be manifest in many ways - such as, for example, leaching from soils, sedimentation in a lake, or the dissolved load in the discharge from a catchment area. Based on the relationship between cyclic and loss processes, the system's ecological efficiency may be assessed. It approaches unity where material cycling is closed (high chemical efficiency) and the damping of the sun's energy pulse is equalised (high thermal efficiency) (Fig 05, 10).
Short-circuited cycling is the result of self-optimisation in the ecosystem - by way of a well-interconnected functioning community of organisms (biocoenosis). Self-optimisation can be achieved only with a system that is limited (space or resource-limited and time or energy-limited). The criterion for optimal integration of organisms is the efficiency: all organisms that contribute to the better governing of closely-coupled cycles help to stabilise the material existence of the ecosystem and thus its sustainability (K-strategy). An ecosystem optimised in such a way can be regarded as an intact system (Fig 11).
If cycles are disturbed, if the losses of a system increase and if the (daily) pulsed solar energy is converted more into heat and accelerated random particle movement, then the system degrades: its intactness is lost. Examples given are the loss of minerals and nutrients from soils and the subsequent material overloading of surface water, where it stimulates eutrophication and the promotion of r-strategists. Stability and sustainability are reduced (see Schrödinger, 1967) (Fig 12).
We have shown that the efficiency principle is applicable at all the various fractal (spatio-temporal) levels of ecosystems. Harmonious and stochastic processes arise in water, which, even at the molecular level, is structured by ions, macromolecules and clusters. The physical and chemical characteristics of water may be understood rather as oscillation or vibration patterns, which are characterised by coherent (i.e. in-phase) resonances, with acceleration or delay of ions and molecules at phase boundaries. During biochemical evolution, these oscillation patterns affected the development of systems through a process of sorting - as stable long-lasting structures would be selected, according to the efficiency principle. These oscillation patterns are the basis of life processes in cells and organisms.
Continuing to higher fractal levels, we have shown that the integration of closely-connected organisms in an ecosystem also accord to the efficiency principle. A simple ecosystem comprises primary producers, the storage/buffer of detritus, consumers, decomposers and water as the transport and reaction medium. The smallest representative and fully functional part of an ecosystem accommodating all five components is called a Dissipative Ecological Unit (DEU). The association of these five functions is necessary to the ability to close a cycle of matter cycles - or, on disturbance, to yield losses. Thus, the DEU, like the ecosystem and landscape of which it is part, is according to the efficiency principle.
The working of the efficiency principle at these higher levels is demonstrated by reference to the intact and degraded (transient) ecosystem 'compartments' of air, vegetation, soil, running waters and lakes. Short-circuited atmospheric-water cycles are characteristic of intact landscape areas carrying vegetation. Since the vegetation develops a powerful humic layer, the water-retention capacity remains high in these areas. Even at times of high temperature, evaporation, and thus the cooling of the land surface, can be maintained. Since larger temperature gradients in the landscape are then much reduced or missing, large spatial climatic events are minimised. Discharges from these intact catchment areas are relatively even and, given good retention capabilities of catchments, are poor in nutrients and mineral ions. Lakes are always subordinate to the functionality of their catchment areas but they will usually develop intact systems under oligotrophic or, at most, mesotrophic conditions. In such lakes, littoral structures, characteristically developing controlled, closed cycles (macrophytes, biofilm), tend to dominate. Production and decompostion are in balance and material losses to the sediments are small. The rate of ageing of an intact lake (i.e. its rate of silting-up) is slowed as long as the functioning of the system continues to self-optimise (Fig 11).
The present impact of humans on landscapes is to degrade their ecosystems in almost every respect. With the removal of vegetation, the cooling function of landscape areas is decreased dramatically and material losses in run-off discharges from catchments are very much increased. Receiving waters become more heavily loaded, so that littoral vegetation and biofilm structures decrease and, in many cases, plankton becomes the dominant controlling component. Through increased sedimentation, the rate of ageing of a lake is accelerated. In addition, lakes may be subject to internal fertilisation from sediments loaded with degradable organic substances, through desulphurisation and ligand exchange of sulphide for phosphate (Fig 12).
Humans do not stand outside the natural system by any means (Fig 16). We still depend on the oxygen in the atmosphere, on a tolerable climate with a generally constant and even precipitation, and on the fertility of soils. Therefore, all management measures and efforts towards reorganisation and restoration of the landscape are subject to exactly the same selection criteria as operate in natural landscapes. With a higher efficiency, the sustainability of the system rises; with high losses, it falls. Without a functional understanding and the adjustment of human activities to conform to the functional conditions of natural systems, extensive breakdowns arising from misuse can hardly be prevented. Against this eventuality, the openness within defined systems should be reduced as far as possible, through intelligent management, where the ageing of the landscape as a whole is slowed down and natural stability is increased. New structures, based on geographical catchments and with humankind integrated as system helmsmen, are necessary. Guided always by the efficiency principle, humankind should be deploying its scientific wherewithal to the development of new, sustainable structures.
Acknowledgements
The basis for this contribution, the ETR model, was developed at the Department of Limnology at the Technical University (Technische Universität) Berlin, under the direction of W. Ripl in cooperation with M. Feibicke, C. Hildmann, K.-D. Wolter and further colleagues. For continuing suggestions and the critical examination of the manuscript we thank I. Otto. Also thanked are S. Ridgill and C.S.Reynolds for improvements to the English language of the text.
References
- Björk, S. (1988) Redevelopment of lake ecosystems - a case study approach. Ambio 17(2), 90-98.
- Brinck, P., Nilsson, L.M. & Svedin, U. (1988) Ecosystem redevelopment. Ambio 17(2), 84-89.
- Cope, F.W. (1972) Structured water and complexed Na+ and K+ in biological systems. In: Water Structure at the Water-Polymer Interface. (ed. H.H.G. Jellinek), pp. 14-18. Plenum Press, New York.
- Cunningham, K.M., Goldberg, M.C. & Weiner, E.R. (1987) An examination of iron oxyhydroxide photochemistry as a possible source of hydroxyl radical in natural waters. In: Chemical Quality of Water and the Hydrologic Cycle. (ed. R.C. Averett & D.M. McKnight), pp. 359-363. Lewis, Chelsea
- Digerfeldt, G. (1972) The post-glacial development of Lake Trummen. Regional vegetation history, water level changes and paleolimnology. Folia Limnologica Scandinavia 16.
- Ebeling, W. & Feistel, R. (1982) Physik der Selbstorganisation und Evolution. Akademie-Verlag, Berlin.
- Eigen, M. & Schuster, P. (1979) The Hypercycle. A Principle of Natural Self-organization. Springer, Berlin.
- Eigen, M. & Winkler, R. (1993) Laws of the Game. How the Principles of Nature Govern Chance. Princeton University, Princeton.
- Gutmann, V. (1978) The Donor-acceptor Approach to Molecular Interactions. Plenum Press, New York.
- Hildmann, C. (1999) Temperaturen in Zönosen als Indikatoren zur Prozeßanalyse und zur Bestimmung des Wirkungsgrades. Energiedissipation und beschleunigte Alterung der Landschaft. Dissertation Technische Universität Berlin, Fachbereich Umwelt und Gesellschaft. D 83. Mensch & Buch, Berlin.
- Hrbácek, J. (1994) 5. Food web relations. In: Restoration of Lake Ecosystems - a Holistic Approach. (ed. M. Eiseltová), pp. 44-58, IWRB Publication 32. International Waterfowl and Wetlands Research Bureau, Slimbridge.
- Lehninger, A.L. (1978) Biochemistry. The Molecular Basis of Cell Structure and Function. Worth Publications, New York.
- Ling, G.N. (1984) In Search of the Physical Basis of Life. Plenum Press, New York.
- Lundqvist, G. (1927) Bodenablagerungen und Entwicklungstypen der Seen. Die Binnengewässer 2, 1-124.
- Mortimer, C.H. (1941/1942) The exchange of dissolved substances between mud and water in lakes. Journal of Ecolology 29, 280-329; 30, 147-201.
- Odum, E.P. (1969) The strategy of ecosystem development. Science 164, 262-70.
- Ohle, W. (1953) Der Vorgang rasanter Seenalterung in Holstein. Naturwissenschaften 40, 153-62.
- Ohle, W. (1954) Sulfat als 'Katalysator' des limnischen Stoffkreislaufes. Vom Wasser 21, 13-32.
- Ohle, W. (1971) Gewässer und Umgebung als ökologische Einheit in ihrer Bedeutung für die Gewässereutrophierung. Wasser - Abwasser (Aachen) 1971(2), 437-56.
- Prigogine, I. (1980) From Being to Becoming. Time and Complexity in the Physical Sciences. Freemann, New York.
- Resch, G. & Gutmann, V. (1987) Wissenschaftliche Grundlagen der Homoeopathie. 2nd edn. Österreichischer Verlag, Berg am Starnberger See.
- Ripl, W. (1978) Oxidation of Lake Sediments with Nitrate - A Restoration Method for Former Recipients. Institute of Limnology, University of Lund.
- Ripl, W. (1992) Management of water cycle: An approach to Urban Ecology. Water Pollution Research Journal of Canada 27(2), 221-37.
- Ripl, W. (1995a) Management of water cycle and energy flow for ecosystem control: the energy-transport-reaction (ETR) model. Ecological Modelling 78, 61-76.
- Ripl, W. (1995b) Der landschaftliche Wirkungsgrad als Maß für die Nachhaltigkeit. In: Umwelt und Fernerkundung: Was leisten integrierte Geo-Daten für die Entwicklung und Umsetzung von Umweltstrategien. (eds R. Backhaus & A. Grunwald), pp. 40-52. Wichmann, Heidelberg.
- Ripl, W. & Hildmann, C. (1997): Ökosysteme als thermodynamische Notwendigkeit. In: Handbuch der Umweltwissenschaften. (eds O. Fränzle, F. Müller & W. Schröder), Ch. V-3.1.1. Loseblattsammlung . Ecomed, Landsberg am Lech.
- Ripl, W. & Hildmann, C. (2000) Dissolved load transported by rivers as an indicator of landscape sustainability. Ecological Engineering 14, 373-87.
- Ripl, W., Hildmann, C., Janssen, T., Gerlach, I., Heller, S. & Ridgill, S. (1995) Sustainable redevelopment of a river and its catchment - the Stör River Project. In: Restoration of Stream Ecosystems. An Integrated Catchment Approach, pp. 76-112. (eds M. Eiseltová & J. Biggs), IWRB Publication 37. International Waterfowl and Wetlands Research Bureau, Slimbridge.
- Ripl, W., Janssen, T., Hildmann, C. & Otto, I. (eds) (1996) Entwicklung eines Land-Gewässer Bewirtschaftungskonzeptes zur Senkung von Stoffverlusten an Gewässer (Stör-Projekt I und II) (in conjunction with F. Trillitzsch, R. Backhaus, H.-P. Blume, P. Widmoser (eds). Im Auftrag des Bundesministeriums für Bildung, Wissenschaft, Forschung und Technologie (BMBF) und des Landesamtes für Wasserhaushalt und Küsten Schleswig-Holstein. Endbericht.
- Ripl, W. & Lindmark, G. (1979) The impact of algae and nutrient composition on sediment exchange dynamics. Archives of Hydrobiology 86(1), 45-65.
- Schrödinger, E. (1967). What is life? Cambridge University Press, Cambridge.
- Vollenweider, R.A. (1971) Scientific Fundamentals of the Eutrophication of Lakes and Flowing Waters, with Particular Reference to Nitrogen and Phosphorus as Factors in Eutrophication. Organisation for Economic Co-operation and Development (OECD), Paris.
- Wetzel, R.G. (1983) Limnology. Saunders, Philadelphia.
- Zachmann, G. (1976) Lexikothek. Spektrum der Naturwissenschaften. Gütersloh.