| Back to 2nd page (with synopsis) |
p.
05 of 11
|
|||
|
|
||||
| Back to last page |
Next
page
|
|
||
There are two focal points of the vegetational distribution of the 2nd transect, stretching through the inner settlement area and broad periphery with individual gardens and fields to the peripheral Macchie zone. The first of these is characterised by the occurrence of ruderal and segetal vegetation in the settlement itself, and the second by the re-occurrence of Macchie vegetation, including the CaS of the extensive pastures and Macchie clearings of the wide village periphery.
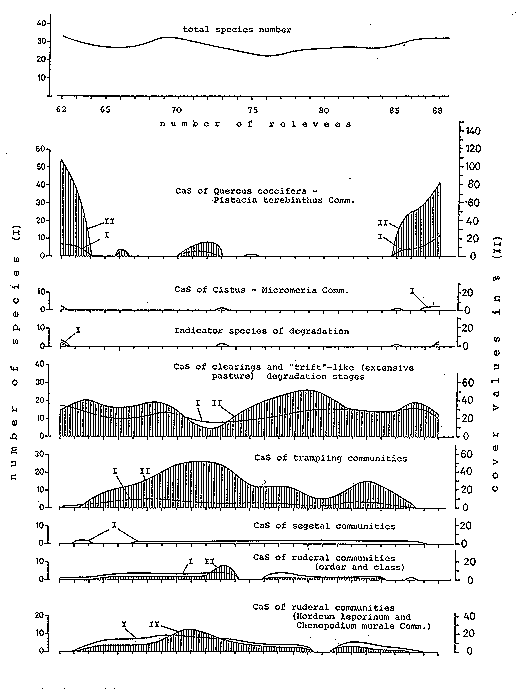
Fig. 6: Distribution of character species groups on the 3rd transect
As already mentioned, the CaS of the extensive pastures are not completely absent even in the centre of the village (3rd transect, cf. Fig. 6), in particular Poa bulbosa and Psilurus incurvus. (Although these were not found in the dry spring of 1979, they were present in almost all soil samples taken from the relevant survey areas for seed bank investigation [cf. 4.6]).
To the north of the highroad around the cistern (see Fig. 4) the species of the extensive pastures increase rapidly, though without reaching the diversity and cover of the first transect. The vegetation of some of the open spaces of the settlement is determined above all by the endemic Verbascum leptocladum with its hairy white leaves. Plantago lagopus and PI. cretica are also well represented along the waysides.
The CaS of the trampling communities, which frequently accompany the segetal, ruderal and Cistus-Micromeria communities (Braun-Blanquet et al. 1951: 128, Oberdorfer 1954: 91, Lohmeyer 1975: 135, Raus 1979: 54, Braun-Blanquet 1964: 498) show a similar distribution in the inner settlement area as the CaS of the ruderal communities. The species abundance varies only slightly, and the highest cover values occurred at the centre of the village. In contrast to the CaS of the trampling communities, the CaS of the ruderal communities intrude much further into the peripheral settlement zones where, together with the CaS of the extensive pasture and Macchie clearings, particularly large numbers of species occur. There is a marked difference here between the distribution of the CaS of the Hordeum leporinum (HI) and the Chenopodium murale (Cm) communities (cf. Table 2). In the village centre and in the northern section around the building complexes, the CaS of the HI community, which is well represented over the whole of the second transect, has higher abundance and dominance. In contrast, the CaS of the Cm community are clearly concen-trated in the inner settlement area, where they establish particularly high dominance to-gether with the CaS of the HI community.
The distribution of the CaS of the segetal vegetation (field weeds) is dependent on the margins of the cornfields. Such species occurring primarily in fields, along the walls (cf. Fig. 11), show an increase in abundance between relevees 25-30 and 43-53, though with only low dominance.
On the 3rd transect, stretching from west to east, evidence of the peripheral village vegetation with abrupt transition to Pinus brutia forest is only provided at a few of the relevees. Although the abundance on the eastern settlement margin is much higher than it is to the west, due to the greater diversity of the Macchie undergrowth, the dominance of Pinus brutia and Quercus coccifera is almost identical. Relics of recurrent bush complexes with Quercus coccifera, Ephedra fragilis and Asparagus acutifolius between the edge of the fields and the dry stone walls can also be found in the village.
Only a few of the CaS of the Phrygana occur together with the CaS of the Macchie. As on the 2nd transect, on the 3rd transect Teucrium polium is the most common. Cistus creticus and Fumana thymifolia occur sporadically, solely on the western forest edge where the undergrowth is sparser.
The CaS of the extensive pastures and Macchie clearings have a very regular distribution. The slight decrease in the numbers of species in the inner settlement area is accompanied by a sharp decline in the dominance values.
Abundance and cover of the broad settlement margin are similar to those of the 2nd transect. In particular there are marked similarities between the northern section of the 2nd transect and the eastern section of the 3rd transect, with intermittent buildings along the paths or road. A somewhat higher cover and a slightly increased abundance at the very edge of the inner settlement area is a common feature of both transect sections. Although the characteristic species of the extensive pastures and their associations are represented almost throughout the transect, they do show considerably different abundance and above all cover values. Of all the species identified in this group, Cynodon dactylon has the highest cover values, and reaches about 60% with the other character species of the extensive pasture associations, which occur mainly in the village centre. The further slight increase in the eastern regions lies near a number of livestock sheds. In P. brutia stands with thick under-growth the species of the extensive pasture association are absent, with one exception. Cynodon dactylon occurs even in stands with sparse undergrowth. The CaS of the segetal associations are represented by few taxa over the transect, and have very low cover values.
As expected, abundance and cover values of the CaS of the ruderal associations are greatest in the central settlement area. The continually increasing numbers of species towards the centre of the village is similar to the 2nd transect with respect to the percentages of the CaS of the HI- and Cm-associations. The CaS of the Hl-associations are represented relatively uniformly along the 3rd transect, whereas the CaS of the Cm-association are mainly encountered at the centre of the village. The highest cover values of both associations are at the very centre of the village, with a greater proportion of CaS of the Hl-association.
Representing 26%, 11 species belong to the CaS of the extensive pastures and Macchie clearings (with companions) (cf. Table 4). The CaS of the ruderal and segetal associations demonstrate a preponderance of CaS of the ruderal associations, and this in turn of the CaS of the Hordeum leporinum association.
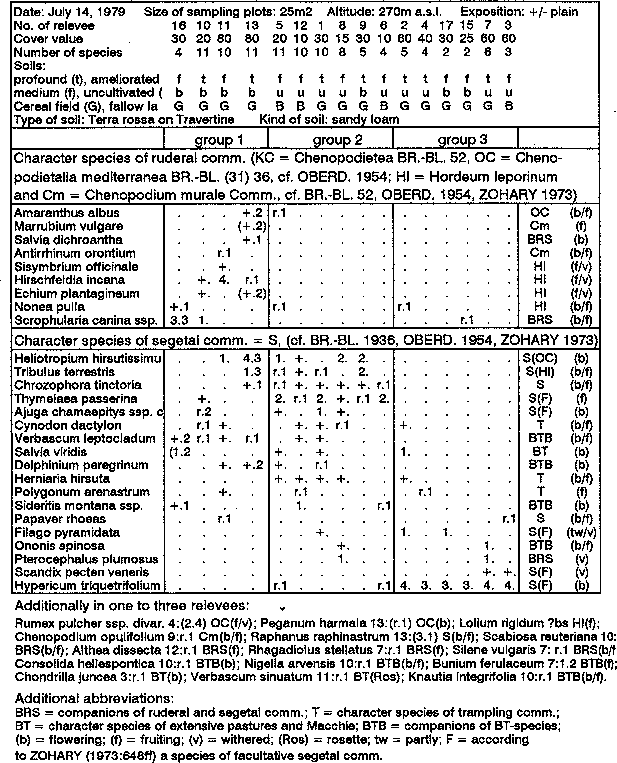
The summary of the vegetation relevees in Table 5 shows the formation of 2 or 3 relevee groups. There is a clear distinction between the virtually monotypic Hypericum group (group 3) with few companions and high cover of Hypericum triquetrifolium, and two groups dominated by the CaS of the ruderal and segetal associations (groups 1 and 2). In addition to a number of 'obligatory' and 'facultative' segetal species (cf. Zohary 1973: 648S), above all Chrozophora tinctoria and Thymelaea passerina, it is only the second group with Heliotropium hirsutissimum and Tribulus terrestris that CaS of Chenopodietalia mediterranea Br.-Bl. (1931) 1936 or of Secalinetalia mediterranea occur with a high degree of regularity (cf. Oberdorfer 1954b and Zohary 1973).
Group 1 differs mainly from those mentioned by the absence of Hypericum triquetrifolium and only a sporadic presence of other segetal species, namely Chrozophora tinctoria and Thymelaea passerina, as well as the increased presence of the character species of the ruderal associations. Dominated by Hypericum spec., the third group is encountered mainly on deep unworked fields or shallow worked fields, as well as (with one exception) in the eastern reaches of the settlement.
On shallower unworked cornfields or fallows with consolidated upper soil horizons in the sparsely inhabited northern part of the settlement, the second CaS group of the ruderal and segetal associations was identified. Two exceptions are no. 8 and 9, which are shallow worked fields.
The first group of the vegetation relevees is concentrated mainly near larger building complexes, or in what is presumably the oldest settlement area. The soils are either deep, or shallow and worked. The latter is the case for no.11, which lies not far from a flat rubbish heap and is dominated by Hirschfeldia incana. The distribution of the CaS of ruderal and segetal associations overlap to a large extent in groups 1 and 2. The character species of the ruderal association are linked more to the centre of the settlement, whereas the segetal species stretch across the middle sections out into the groups 1 and 2.
|
|||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||
The vegetation mapped at the old centre of the settlement near the wall between the open space and the harvested cornfield is dominated by Hirschfeldia incana, a widely spread CaS along paths and edges of fields during summer in the Eastern Mediterranean (Oberdorfer 1954b, Zohary 1973). The relict vegetation of the Macchie is severely devastated (cf. 4.4), but nevertheless extends with some species of the extensive pastures and Macchie clearings into the centre of the village.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||